
Trabalho avalia os efeitos indiretos da herbivoria sobre a fotossíntese e identifica quatro mecanismos que contribuem para a supressão indireta da fotossíntese em tecidos foliares remanescentes: sistema vascular danificado, demanda energética alterada, autotoxicidade induzida por mecanismos de defesa e defesa induzida a regulação negativa da fotossíntese.
O artigo tem autoria de Paul D. Nabity, Jorge A. Zavala e Evan H. DeLucia foi publicado no Ann Bot . 2009 fev; 103 (4): 655-663 e disponibilizado online em 26 de julho de 2008, confira na íntegra acessando aqui.
Os insetos consomem cerca de 14% do total da produção agrícola global, este percentual já considera a aplicação do controle fitossanitário, pois sem o devido controle os danos seriam superiores a 50% para as principais culturas agrícolas (Oerke e Dehne, 1997).
Como o dano foliar é avaliado em lavouras?
Tradicionalmente a quantificação de dano é feita através de um levantamento da quantidade de tecido removido da folhagem, assumindo que, o tecido foliar remanescente funciona normalmente. É comum os níveis de controle assumirem como premissa um percentual de área foliar consumida. Contudo, existe uma série de dano decorrentes do ataque dos insetos que afetam a fotossíntese em tecidos não danificados, e esses efeitos “indiretos” na fotossíntese podem ser consideravelmente maiores do que a remoção direta da área foliar (Welter, 1989 ; Zangerl et al., 2002 ), ou seja, a eficiência do tecido foliar remanescente é alterada devido aos danos ou a ausência do tecido consumido.
São mencionados neste trabalho diferentes tipos de de respostas/prejuízos em decorrência da alimentação dos insetos, vamos especificá-los:
Os efeitos indiretos da herbivoria foram atribuídos a quatro classes: danos ao sistema vascular, demanda energética alterada, autotoxicidade relacionada aos mecanismos de defesa e regulação negativa da fotossíntese induzida pela defesa.

Adaptado do original

Segundo publicação de IOWA STATE UNIVERSITY, o trabalho realizado por Pedigo et al. (1986) (Confira o trabalho na íntegra acessando aqui), demonstra a relação rendimento x lesão em uma curva didática. Nela algumas plantas podem ser capazes de sofrer danos sem afetar a produtividade (tolerância), enquanto algumas, como a soja, podem produzir um rendimento ligeiramente maior com uma pequena quantidade de lesão (supercompensação) mas de maneira geral, incrementando-se as injurias tem-se a perda de rendimento. Veja a curva de dano:
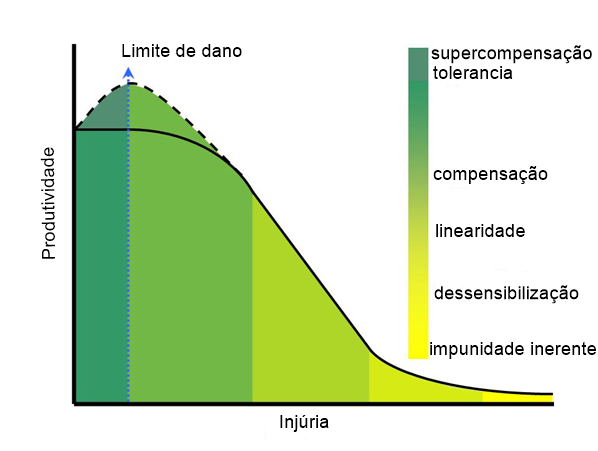
Adaptado de: IOWA
A linha azul tracejada através denota o ponto em que a perda de rendimento é detectável e atribuída à lesão (limite de dano). É importante ressaltar que a função da curva de dano pode mudar dependendo de fatores como o estágio da praga ou planta ou o ambiente. Determinar essa função para uma praga é um passo crítico no desenvolvimento e refinamento de um nível de dano econômico.
Como os métodos amostrais por imagem pode ajudar na mensuração do dano por herbivoria dos insetos?
O uso da análise de imagens geográficas já são utilizadas como ferramenta para fazer comparações quantitativas de imagens, já que esse método permite compilar muitas camadas de informações de co-variáveis para analisar um todo.
A fluorescência da clorofila fornece subsidio para quantificar os processos de componentes relacionados ao transporte de elétrons fotossintéticos, na medida em que as limitações de água ou outras alterações localizadas na química das folhas afetam a condutância estomática tem se variações nas imagens (Jones, 1999 ; Omasa e Takayama, 2003 ).
Com a calibração adequada, os mapas térmicos podem ser convertidos diretamente em mapas de condutância estomática ( Jones, 2004 ; Bajons et al. , 2005 ; Grant et al. , 2006 ), embora seja desafiador devido a divergência de resolução em alguns equipamentos.
Dependendo da intensidade dos danos, ignorar a supressão indireta da fotossíntese por artrópodes e outros organismos pode levar a uma subestimação de seus impactos fisiológicos e ecológicos. Quantificar o dano muitas vezes perpassa a extensão do mesmo, e sim a localização e distribuição nas plantas.
Todas as referencias utilizadas no corpo desta nota técnica estão disponíveis no trabalho original. As demais citações são mencionadas no próprio texto.
Fonte Mais Soja
Oferecimento:





















